А.А. Галанин
Лихенометрический метод датирования: Идентификация и оценка возраста процессов современной морфодинамики в горах Северо-Востока России
СВКНИИ ДВО РАН. 2000. URL: http://academnet.neisri.ru/academnet/neisri/lab/neotecton/lihen/lichenometry2.htm
Содержание раздела
ЛихенометрияВзаимосвязь эпилитных лишайников и литоосновы |  Молодая колония Xanthoria elegans на техногенной кладке из "дикого" камня. Возраст строения 53 года. 24-км Иультинской трассы, хребет Искатень, Чукосткий п-ов |
Лихенометрия
Лишайники
 |
Скорости роста разных лишайников существенно различаются. Относительно быстрорастущими являются некоторые тропические кустистые виды, ежегодный прирост которых достигает более 10 см. Намного медленнее растут тонкие накипные циркумарктические виды, которые наиболее часто используются в лихенометрии, например роды Rhizocarpon, Aspicilia, Leucanora, и др. Скорости роста особей одних и тех же видов зависят от конкретных физико-географических условий и возраста самих особей. |
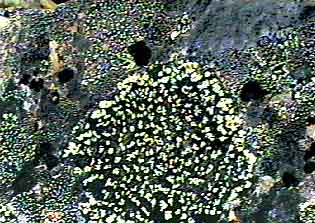
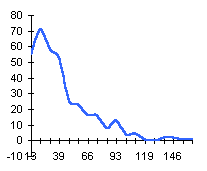 | 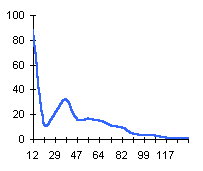 | 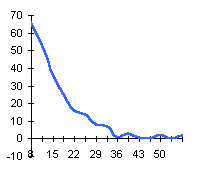 |
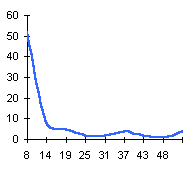 | 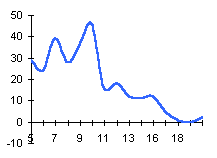 | Частотные распределения лишайников индикаторов на морфоскульптурных поверхностях разного генезиса и современной динамики . По оси X — интервал группирования; по оси Y — диаметры талломов в мм |
где t — минимальный возраст поверхности, dRh — диаметр наиболее крупной особи Rhizocarpon sp. Для лихенометрических оценок Дж. Метьюс [Matthews, 1974; 1975; 1977] вывел уравнение, использующее логарифмическую функцию log(t+c)=a+bx, где t — возраст поверхности, | 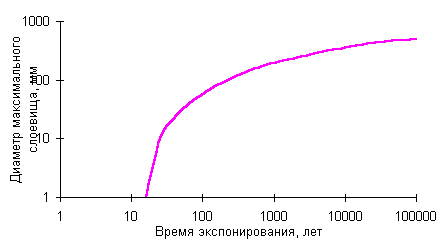
|
лишайника-индикатора, а, b и с — коэффициенты, зависящие от особенностей используемого вида и физико-климатических условий района датирования. Очевидно, что для нахождения этих коэффициентов необходимо решить систему из трех уравнений. Для территории Норвегии этим исследователем коэффициенты а, b и с были оценены в 1,8143, 0,0062 и 50 соответственно. В предлагаемой автором зависимости не возраст, а логарифм возраста линейно связан с диаметром максимального лишайника. Выполненный нами анализ данной зависимости при разных значениях x показал, что она отражает представления Р.Е. Бешеля о фазе быстрого нелинейного роста и, следующей за ней линейной фазе. Однако, по данной функции в интервале возрастов более 100 лет, она принимает форму, близкую к линейной. Что противоречит условию об предельных размерах роста слоевищ накипных лишайников. А.А.Никоновым с соавторами для построения вероятностной модели роста локальных популяций лишайника индикатора использовался известный закон замедления роста численности живых организмов — Т.Р. Мальтуса. Профессионально математически постороенная авторами модель весьма сложна для практического использования, к тому же в ее основе приняты некоторые априорные допущения, противоречащие законам конкуренции и популяционно-ценотических отношений в локальных сообществах накипных лишайников [Галанин, 1997; 1999]. Тем не менее, среди известных лихенометрических методик она является наиболее удовлетворительной, позволяя рассчитать ошибки возрастных оценок.
а после интегрирования Параметры роста в разных районах неодинаковы и зависят от их физико-географических условий, определяющих продолжительность вегетационного периода лишайников. Для субарктической тундры побережья Анадырского лимана параметры ао и f для среднего значения ста максимальных особей соответственно составляют: секция Rhizocarpon — 0,14±0,01 мм/год и 526±95 лет, род Aspicilia — 0,15±0,01 мм/год и 733±115 лет. В лесотундровой зоне центральной части Корякского нагорья для секции Rhizocarpon — 0,18±0,03 мм/год и 452±88 лет, а начальный годовой прирост листоватого лишайника Parmelia sp. — 0,47±0,08 мм/год. В Примагаданье (зона северной тайги) параметры роста лишайников рода Rhizocarpon секции Rhizocarpon составляют 0,23±0,03 мм/год и 419±86 лет. Таким образом, скорости роста исследуемых лишайников в разных физико-географических районах существенно различаются. Наиболее низкие годовые приросты и, соответственно, более медленный рост и развитие популяций накипных лишайников характерно для побережья Анадырского лимана, что не противоречит условиям субарктического климата. Параметры роста накипных лишайников в Северной Корякии и Примагаданье близки по значениям, что, по-видимому, можно объяснить близкими климатическими характеристиками этих районов. |
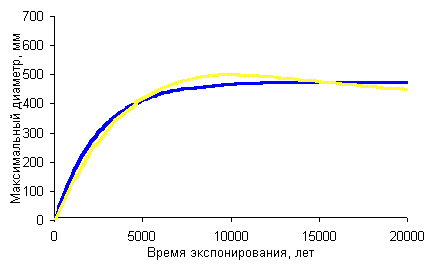 Синяя линия — кривая роста Rhizocarpon geographicum для |
датирования по этой методике составляет около 20%.
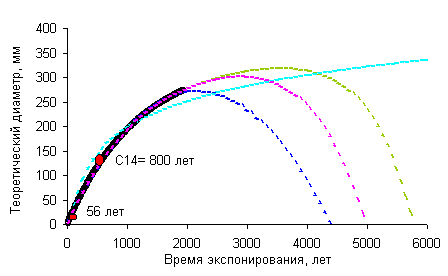 Кривая роста Rhizocarpon sp. для гипсометрического интервала 200-400 м Северо-охотского побережья. Аппроксимационные |
Построение кривых роста с учетом старения локальных лишайниковых популяцийНаблюдения за популяциями эпилитных лишайников показали, что наиболее крупные особи обитают не на самых древних плакорных поверхностях, а на субстрате более молодого возраста. Наиболее крупные лишайники, обитающие на поверхностях, время экспонирования которых превышает этот возрастной оптимум, имеют меньшие диаметры [Галанин, 1996, 1999]. Это объясняется постепенными сукцессионными сменами, замещением эдификаторов и доминантов в локальном сообществе по мере выветривания и разрушения поверхности морфоскульптуры, изменения морфологических и химических свойств субстрата. Поэтому истинная кривая роста наиболее часто используемых эпилитных видов, например, из рода Rhizocarpon, должна описываться полиномиальной зависимостью и иметь математический экстремум, отвечающий субстратно-возрастному оптимуму. |
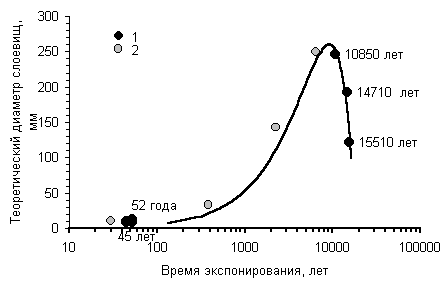 Кривая роста Rhizocarpon sp. для нивально-гляциальных районов |
Взаимосвязь эпилитных лишайников и литоосновыВлияние динамики субстрата на динамику лишайниковых сообществ |
 Сукцессионная стадия замещения лишайников Rhizocarpon sp. на Aspicilia sp. (серый) на старом отмершем куруме (хр. Пекульней). На талломе Aspicilia sp. уже поселилась особь |
Через 30-50 лет от начала колонизации поверхности общий проективный покров лишайников уже достигает 60% и более. Начинается жесткая конкуренция и уменьшение числа особей на каждом локальном участке. Стремясь обеспечить себе необходимое жизненное пространство, особи лишайников как бы “разгоняются” в вершины сетки из более или менее равносторонних треугольников. По-видимому, в результате нарастания на соседние слоевища, доминантная особь частично использует вещество угнетаемого таллома, в результате чего скорость ее роста несколько убыстряется. |
| При этом особи-доминанты чаще всего оказываются в центре шестигранников, сложенных из первоначальных треугольников. Начинает формироваться новый модальный класс особей лидеров, которые также “стремятся разойтись” в вершины треугольников, но уже большего порядка — треугольников, составленных особями, расположенными в центрах первоначальных шестигранников. |  Фрагмент таллома с апотециями циркумполярно распространенного коркового лишайникаHaematomma ventosum (Гематома ветровая). Этот вид наиболее конкурентоспособен на старых плакорных поверхностях, где является эдификатором в сообществах накипных лишайников (увел. в 2 раза) |
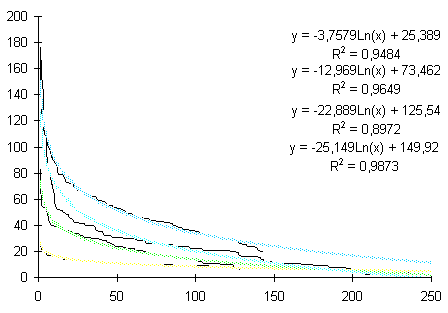
Сукцессионные смены | 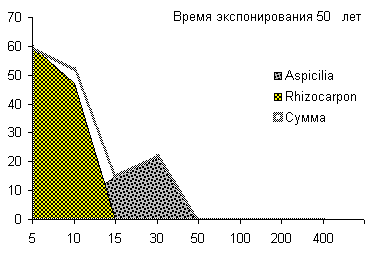 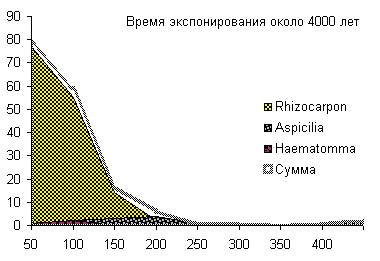 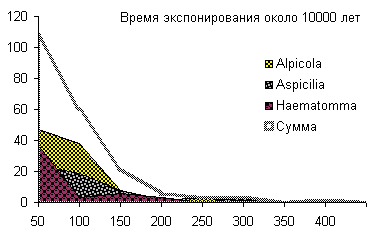 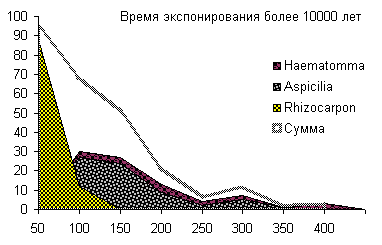 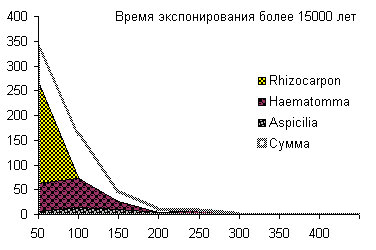 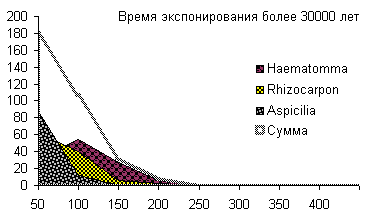 Совмещенные частотные распределения диаметров лишайников различных морфологических групп, являющихся эдификаторами на разных сукцессионных стадиях на каменных поверхностях разной степени выветрелости. По оси X — интервал группирования, мм; по оси Y — частость |
Документ изменен: 01/14/2008 16:07:54
Галанин А.А. Лихенометрический метод датирования: Идентификация и оценка возраста процессов современной морфодинамики в горах Северо-Востока России. СВКНИИ ДВО РАН. 2000
| ЗНа уровень вверх | ЫПредыдущий раздел | ЬСледующий раздел |
| © Copyright A.A.Galanin, North-East Interdisciplinary Scientific Research Institute, 2001. All rights reserved. | Desine by A.A.Galanin. Magadan, 2001
|
Приложение
Измерение скорости роста эпилитных лишайников в поселке Соловецкий
URL: http://lichenhouse.narod.ru/29/2/3.htm
.png)
